标签:
单词:
polymerase聚合酶[‘p?l?m?re?z; p?‘l?m?re?z]
an enzyme that catalyzes(催化) the formation of new DNA and RNA from an existing strand of DNA or RNA
covalently共价的
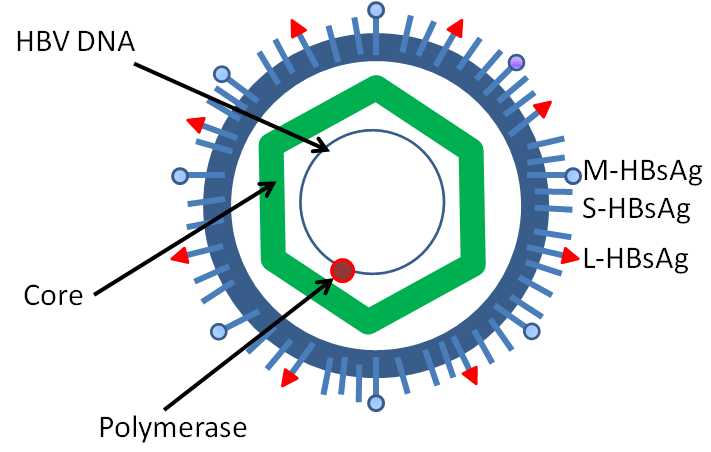
The structure of hepatitis B virus
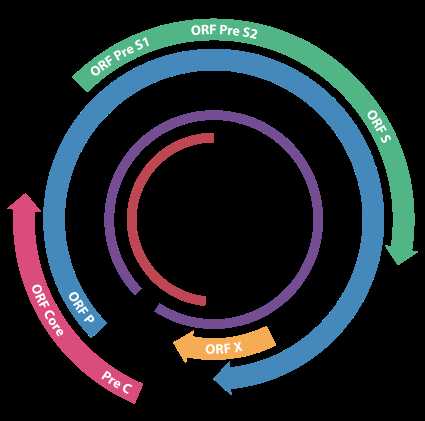
The genome organisation of HBV. The genes overlap.
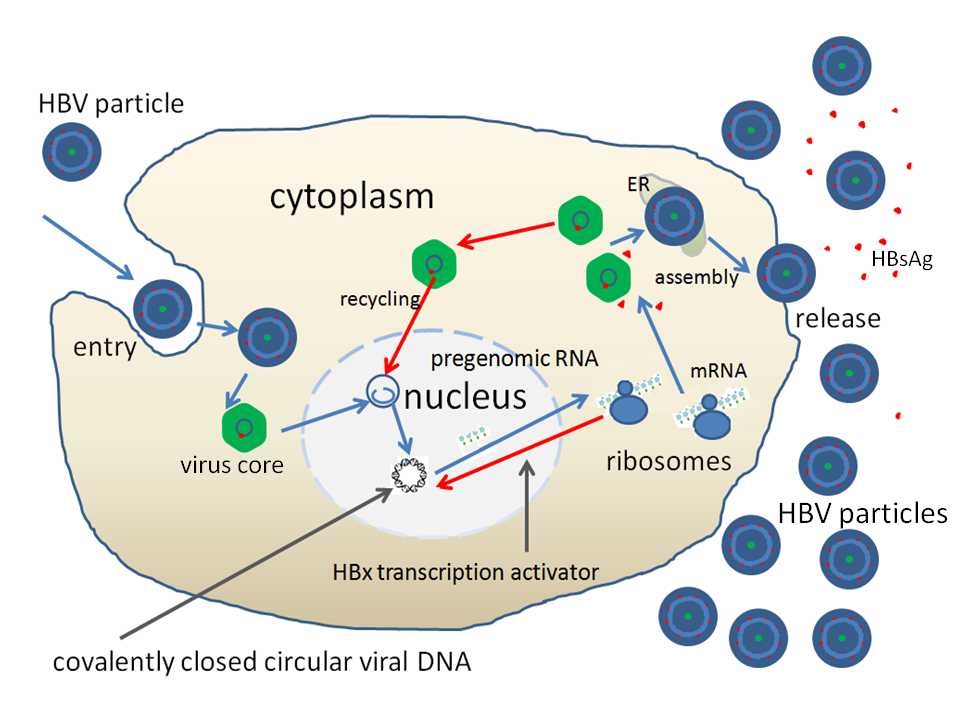
Hepatitis B virus replication
乙型肝炎病毒 (HBV) 属嗜肝DNA 病毒科 (hepadnaviridae),基因组长约3.2kb ,为部分双链环状DNA。
HBV 侵入人体后,与肝细胞膜上的受体结合,脱去包膜,穿入肝细胞质内,然后脱去衣壳,部分双链环状HBV DNA 进入肝细胞核内,在宿主酶的作用下,以负链DNA 为模板延长正链,修补正链中的裂隙区,形成共价闭合环状DNA (cccDNA) ,然后以cccDNA 为模板,在宿主RNA 聚合酶II 的作用下,转录成几种不同长短的mRNA ,其中3.5kb 的mRNA 含有HBV DNA 序列上全部遗传信息,称为前基因组RNA。后者进入肝细胞质作为模板,在HBV 逆转录酶作用下,合成负链DNA; 再以负链DNA 为模板,在HBV DNA 聚合酶作用下,合成正链DNA,形成子代的部分双链环状DNA, 最后装配成完整的HBV,释放至肝细胞外。胞质中的子代部分双链环状DNA 也可进入肝细胞核内,再形成cccDNA 并继续复制。cccDNA 半寿 (衰) 期长,很难从体内彻底清除[1, 2]。
HBV 含4个部分重叠的开放读码框 (ORF),即前S/S 区、前C/C 区、P 区和X 区。前S/S 区编码大 (前S1、前S2 及S)、中 (前S2 及S)、小 (S) 3 种包膜蛋白;前C/C 区编码HBeAg 及HBcAg;P 区编码聚合酶;X 区编码X 蛋白。
前C 区和基本核心启动子 (BCP) 的变异可产生HBeAg 阴性变异株。前C 区最常见的变异为G1896A 点突变,形成终止密码子 (TAG) ,不表达HBeAg。BCP 区最常见的变异是A1762T/G1764A 联合点突变,选择性地抑制前C mRNA 的转录,降低HBeAg 合成[3]。
P 基因变异主要见于POL/RT 基因片段 (349~692 aa ,即rt1~rt344) 。在拉米夫定治疗中,最常见的是酪氨酸-蛋氨酸-天门冬氨酸-天门冬氨酸 (YMDD) 变异,即由YMDD 变异为YIDD (rtM204I) 或YVDD (rtM204V),并常伴有rtL180M 变异,且受药物选择而逐渐成为对拉米夫定耐药的优势株[4] (Ⅰ)。
S 基因变异可导致隐匿性HBV 感染(occult HBV infection) ,表现为血清HBsAg 阴性,但仍可有HBV 低水平复制 (血清HBV DNA 常 < 104 拷贝/ml)[5]。
根据HBV 全基因序列差异≥8%或S 区基因序列差异≥4%,目前HBV 分为A~H 8 个基因型。各基因型又可分为不同基因亚型。A 基因型慢性乙型肝炎患者对干扰素治疗的应答率高于D 基因型,B 基因型高于C 基因型;A 和D 基因型又高于B 和C 基因型[6] (Ⅰ)。基因型是否影响核苷 (酸) 类似物的疗效尚未确定。
HBV 易发生变异。在HBV 感染者体内,常形成以一个优势株为主的相关突变株病毒群,称为准种 (quasispecies),其确切的临床意义有待进一步证实。
HBV 的抵抗力较强,但65℃10 h 、煮沸10 min 或高压蒸气均可灭活HBV。含氯制剂、环氧乙烷、戊二醛、过氧乙酸和碘伏等也有较好的灭活效果。
标签:
原文地址:http://www.cnblogs.com/biopy/p/4337168.html